umat Kristiani khususnya Katolik di Kota Kupang yang ingin mengadakan
wisata rohani pada umumnya akan memilih Larantuka, Bitauni atau bagi
yang mampu akan memilih Timur Tengah, Fatima (Portugal), atau Lourdez
(Perancis). Namun di Wilayah Kabupaten Kupang yang tidak jauh dari Kota
Kupang, ada kawasan taman doa yang tidak kalah indah bagi mereka yang
menyukai keunikan alam maupun bagi orang2 yang ingin khusyuk beribadah.
lebih lengkapnya dapat dibaca di sini
Rabu, 06 Juni 2012
Selasa, 05 Juni 2012
PROTEIN

Fungsi Protein
- Pembentukan dan pemeliharaan tubuh
Sebelum sel-sel dapat mensintesis protein baru, harus tersedia
semua asam amino esensial yang diperlukan dan cukup nitrogen atau ikatan amino
(NHz) guna pembentukan asam-asam amino nonesensial yang diperlukan. Pertumbuhan
at au penambahan otot hanya mungkin bila tersedia cukup campuran asam amino
yang sesuai termasuk untuk pemeliharaan dan perbaikan. Beberapa jenis jaringan tubuh membutuhkan
asam-asam amino, tertentu dalam jumlah lebih besar. Rambut, kulit, dan kuku
membutuhkan lebih banyak asam amino yang mengandung sulfur. Protein kolagen merupakan protein utama
otot urat-urat dan jaringan ikat. Fibrin dan miosin adalah protein lain yang
terdapat di dalam otot-otot.
Protein
tubuh berada dalam keadaan dinamis, yang secara bergantian dipecah dan disintesis kembali. Tiap
hari sebanyak 3 % jumlah protein total berada dalam keadan berubah ini. Dinding
usus yang setiap 4-6 hari harus diganti, membutuhkan sintesis 70 gram protein
setiap hari. Tubuh sangat efisien dalam memelihara protein yang ada dan
menggunakan kembali asam amino yang diperoleh dari pemecahan jaringan untuk
membangun kembali jaringan yang sarna atau jaringan lain.
- Pembentukan ikatan – ikatan esensial tubuh
Hormon-hormon, seperti tiroid, insulin, dan epinefrin
adalah protein, demikian pula berbagai enzim. Ikatan-ikatan ini bertindak
sebagai katalisator at au membantu perubahan-perubahan biokimia yang terjadi di
dalam tubuh.
Hemoglobin,
pigmen darah yang berwarna merah dan berfungsi sebagai pengangkut oksigen dan
karbon dioksida adalah ikatan protein. Begitupun bahan-bahan lain yang berperan dalam penggumpalan darah. Protein lain adalah
fotoreseptor pada mata.
Asam amino
triptofan berfungsi sebagai prekursor vitamin niasin dan pengantar saraf
serotonin yang berperan dalam membawa pesan dari sel saraf yang satu ke yang
lain.
Dalam hal kekurangan protein, tampaknya tubuh
memperioritaskan pembentukan ikatan-ikatan tubuh yang vital ini.
- Mengatur keseimbangan air
Cairan tubuh terdapat di dalam tiga kompartemen:
intraselular (di dalam sel), ekstraselular/interselular (di antara sel), dan
intravaskular (di dalam pembuluh darah). Kompartemen-kompartemen ini dipisahkan satu sama lain oleh membran sel.
Distribusi cairan di dalam kompartemen-kompartemen ini harus dijaga dalam
keadaan seimbang atau homeostasis. Keseimbangan ini diperoleh melalui sistem
kompleks yang melibatkan protein dan elektrolit. Penumpukan cairan di dalam
jaringan dinamakan edema dan merupakan tanda awal kekurangan protein.
- Memelihara netralitas tubuh
Protein tubuh bertindak sebagai buffer, yaitu bereaksi
dengan asam dan basa untuk menjagaa pH pada taraf konstan. Sebagian besar
jaringan tubuh berfungsi dalam keadaan pH netral atau sedikit alkali (pH 7,35 -
7,45).
- Pembentukan antibodi
Kemampuan tubuh untuk memerangi infeksi bergantung pada
kemampuannya untuk memproduksi antibodi terhadap organisme yang menyebabkan
infeksi tertentu atau terhadap bahan-bahan asing yang memasuki tubuh. Tingginya
tingkat kematian pada anak-anak yang menderita gizi-kurang kebanyakan
disebabkan oleh menurunnya daya tahan terhadap infeksi (muntaber, dan
sebagainya) karenaketidakmampuannya membentuk antibodi dalam jumlah yang cukup.
Kemampuan tubuh untuk melakukan detoksifikasi terhadap
bahan-bahan racun dikontrol oleh enzim-enzim yang tertutama terdapat di dalam
hati. Dalam keadaan kekurangan protein kemampuan tubuh untuk menghalangi
pengaruh toksik bahan-bahan racun ini berkurang. Seseorang yang menderita kekurangan protein lebih
rentan terhadap bahan-bahan racun dan obat-obatan.
- Mengangkut zat – zat Gizi
Protein
memegang peranan esensial dalam mengangkut zat-zat gizi dari saluran cerna
melalui dinding saluran cerna ke dalam darah, dari darah ke
jaringan-jaringan, dan melalui membran sel ke dalam sel-sel. Sebagian besar
bahan yang mengangkut zat-zat gizi ini adalah protein. Alat angkut protein ini
dapat bertindak secara khusus, misalnya protein pengikat-retinol yang hanya
mengangkut vitamin A. Atau dapat mengangkut beberapa jenis zat gizi seperti
mangan dan zat besi, yaitu transferin; atau mengangkut lipida dan bahan
sejenis-lipida, yaitu lipoprotein.
Kekurangan
protein, menyebabkan gangguan pada absorpsi dan transportasi zat-zat gizi.
- Sumber energi
Sebagai
sumber energi,
protein ekivalen dengan karbohidrat, karena menghasilkan 4 kkal/g protein.
Namun, protein sebagai sumber energi relatif lebih mahal, baik dalam harga
maupun dalam jumlah energi yang dibutuhkan untuk metabolisme energi.
Fungsi Protein sebagai Biomolekul
•
Katalisator
(enzim dan hormon insulin)
•
Molekul karier (hemoglobin, lipoprotein)
•
Reseptor signal biologik
•
Komponen
struktural, kolagen dlm urat dan tulang rawan, elastin pd persendian, keratin
pd kuku, bulu, rambut, fibrous pd laba2 dan kepompong ulat sutra.
Kunjungi pula : http://nurdhienln.wordpress.com
Kunjungi pula : http://nurdhienln.wordpress.com
Senin, 04 Juni 2012
KARBOHIDRAT

Klasifikasi Karbohidrat
1.
Karbohidrat sederhana
•
Monosakarida
tdd: glukosa, fruktosa, galaktosa, manosa dan pentosa
•
Disakarida
tdd: sukrosa (glukosa & fruktosa), maltosa (2 glukosa), laktosa (glukosa
& galaktosa), trehalosa (2 glukosa).
•
Gula
alkohol (bentuk alkohol dr monosakarida) tdd : manitol, sorbitol dan inositol
•
Oligosakarida
tdd: rafinosa, stakiosa, verbaskosa dan fruktan
2.
Karbohidrat kompleks
•
Polisakarida tdd: pati, dekstrin, glikogen
•
Serat
tdd : selulosa, hemiselulosa, lignin (tidak larut dalam air) dan pektin, gum,
mukilase, glukan, algal (larut dalam air)
Fungsi karbohidrat
Sumber energi
Fungsi utama karbohidrat adalah
menyediakan energi bagi tubuh. Karbohidrat merupakan sumber urama energi bagi
penduduk di seluruh dunia, karena banyak di dapat di alam dan harganya relatif
murah. Satu gram karbohidrat menghasilkan 4 kkalori. Sebagian karbohidrat di
dalam tubuh berada dalam slrkulasi darah sebagai glukosa untuk keperluan energi
segera; sebagian disimpan sebagai glikogen dalam hati dan jaringan otot, dan
sebagian diubahmenjadi lemak untuk kemudian disimpan sebagai cadangan energi di
dalam jaringan lemak. Seseorang yang memakan karbohidrat dalam jumlah
berlebihan akan menjadi gemuk. Sistem saraf sentral dan otak sama sekali
tergantung pada glukosa untuk keperluan energinya.
Pemberi rasa manis
Karbohidrat
memberi rasa manis pada makanan, khususnya mono dan disakarida. Sejak lahir
manusia menyukai rasa manis. Alat kecapan pada ujung lidah merasakan rasa manis
tersebut. Gula tidak mempunyai rasa manis yang sama. Fruktosa adalah gula
paling manis. Bila tingkat kemanisan
sakarosa diberi nilai 1, maka tingkat
kemanisan frukrosa adalah 1,7; glukosa
0,7; maltosa 0,4; laktosa 0,2

Penghemat protein
Bila
karbohidrat makanan tidak mencukupi, maka protein akan digunakan untuk memenuhi kebutuhan
energi, dengan mengalahkan fungsi utamanya sebagai zat pembangun. Sebaliknya
bila karbohidrat makanan mencukupi, protein terutama akan digunakan sebagai zat
pembangun.
Pengatur metabolisme lemak
Karbohidrat mencegah terjadinya oksidasi lemak yang tidak
sempurna, sehingga menghasilkan bahan-bahan keton berupa asam asetoasetat, aseton,
dan asam beta-hidroksi-butirat. Bahan-bahan
ini dibentuk dalam hati dan dikeluarkan melalui urine dengan mengikat basa
berupa ion natrium. Hal ini dapat menyebabkan ketidakseimbangan natrium dan
dehidrasi. pH cairan tubuh menurun. Keadaan ini menimbulkan ketosis at au
asidosis yang dapat merugikan tubuh. Dibutuhkan antara 50-100 gram karbohidrat
sehari untuk mencegah ketosis.
Memperlancar pencernaan
Karbohidrat
membantu pengeluaran feses dengan cara mengatur peristaltik usus dan memberi bentuk
pada: feses. Selulosa dalam serat makanan mengatur peristaltik usus, sedangkan
hemiselulosa dan pektin mampu menyerap banyak air dalam usus besar sehingga
memberi bentuk pada sisa makanan yang akan dikeluarkan.
Serat
makanan mencegah kegemukan, konstipasi, hemoroid, penyakit-penyakit divertikulosis,
kanker usus besar, penyakit diabetes mellitus, dan jantung koroner yang
berkaitan dengan kadar kolesterol darah tinggi.
Laktosa
dalam susu membantu absorpsi kalsium. Laktosa lebih lama tinggal dalarr saluran
cerna, sehingga menyebabkan pertumbuhan bakteri yang menguntungkan. Bakteri
tertentu diduga mensintesis vitamin-vitamin tertentu dalam usus besar. Asam
glukoronat turunan glukosa, di dalam hati mengikat toksin-toksin dan bakteri
dan mengubahnya menjadi bentuk-bentuk yang dapat dikeluarkan dari tubuh.
Genetika
Gula
ribosa yang mengandung lima atom karbon merupakan bagian dari ikatan DNA dan
RNA.

PENGANTAR ILMU GIZI KESEHATAN MASYARAKAT
Pendahuluan
Mempelajari tentang gizi kini tidak hanya tentang
zat gizi tersebut yang mencakup bentuk; sifat; dan ketersediaannya, serta
fungsi, kegunaan dan interaksinya dengan jaringan tubuh. Tetapi kini ilmu gizi
dipelajari mencakup seluruh aspek penyediaan makanan dan zat gizi yang mencakup
produksi, distribusi dan ketersediaan pangan, keadaan sosial ekonomi
masyarakat, budaya yang berkaitan dengan makanan serta pengetahuan dan
pemahaman masyarakat tentang makanan. Sebab telah diketahui bahwa konsumsi gizi
seseorang sangat berkaitan dengan hal-hal di atas.
Definisi
Ilmu gizi kesehatan masyarakat adalah salah satu
cabang dari ilmu gizi yang mempelajari tentang gizi pada suatu masyarakat. Dengan
demikian yang dipelajari adalah tentang status konsumsi pangan serta status
gizi masyarakat dan bukan perorangan. Bagian yang paling banyak dipelajari
adalah faktor-faktor yang mempengaruhi keadaan konsumsi dan status gizi
masyarakat.
Hubungan Ilmu Gizi Kesehatan
masyarakat dengan ilmu-ilmu lain.
1. Biologi
Biologi adalah ilmu yang
mempelajari tentang hal-hal yang bersifat hayati. Di dalam biologi dipelajari
tentang manusia, tumbuh-tumbuhan dan hewan yang mencakup jenis dan
sifat-sifatnya. Pengetahuan ini berguna dalam menentukan dan menemukan
sumber-suber makanan serta budidayanya.
Selain itu dipelajari pula
tentang ilmu manusia baik anatomi dan fisiologi sehingga secara tidak langsung
dapat dipelajari aspek dinamis yang dilakukan tubuh terhadap makanan atau yang
dialami makanan yang mencakup pencernaan, absorbsi, distribusi, metabolisme dan
ekskresi.
2. Kimia.
Dalam ilmu kimia dipelajari tentang materi dan
interaksinya. Makanan yang termasuk ke dalam unsur-unsur dan senyawa kimia
turut mengalami reaksi. Reaksi itu mencakup reaksi antar makanan, dengan
lingkungan di luar tubuh dan juga reaksi di dalam tubuh termasuk metabolisme.
3.
Ekonomi
Terdapat hubungan saling mempengaruhi antara
status gizi dengan kondisi perekonomian masyarakat.
Hubungannya berupa hubungan yang timbal balik. Pangan yang tersedia
memungkinkan masyarakat untuk meningkatkan pendapatannya secara langsung
khususnya bagi masyarakat yang bekerja di sektor pertanian. Bahan pangan
sebagai salah satu kebutuhan primer sehingga pergerakannya di pasar tergolong fast moving sehingga meningkatnya
produksi pangan yang beredar di pasar secara tidak langsung meningkatkan status
perekonomian masyarakat di daerah tersebut. Jika produksi pangan meningkat maka
ada kemungkinan tercipta harga yang optimal bagi daya beli masyarakat maka
konsumsi akan meningkat dan dengan tubuh yang sehat, masyarakat akan menjadi
lebih produktif baik secara sosial dan ekonomi.
Pengaruh ekonomi terhadap status gizi yaitu dengan
meningkatnya pendapatan dan daya beli masyarakat maka masyarakat dapat dengan
leluasa memenuhi kebutuhan pangan khususnya untuk ketersediaan pangan tingkat
rumah tangga.
Itulah sebabnya dalam mempelajari status gizi
masyarakat maka harus dipelajari pula status ekonomi masyarakat tersebut yang
mencakup jenis mata pencaharian dan pendapatan.
4. Geografi
Dalam geografi dipelajari tentang geologi, hidrologi,
klimatologi dan demografi. Geologi mempelajari tentang struktur permukaan bumi
apakah itu berupa dataran rendah, tinggi, atau parmukaan curam. Selain itu
dipelajari pula struktur tanah dan batuan apakah tipe mediteran, aluvial dan
lain-lain. Sehingga dengan mempelajari hal-hal tersebut dapat diketahui
jenis-jenis tanaman pangan yang bisa tumbuh di suatu wilayah.
Dalam hidrologi dipelajari tentang air yang
terdapat di alam, dan Klimatologi mempelajari tentang iklim dan cuaca. Sehingga
bisa diketahui tempat dan saat yang tepat di mana kondisi kelembaban dan
ketersediaan air tanah dan air permukaan serta kondisi cuaca yang sesuai untuk
tanaman pangan.
Dalam demografi mempelajari tentang karakteristik
penduduk yang mencakup jumlah, kepadatan, mata pencaharian, pendapatan dan
lainnya sehingga dapat diketahui kondisi ketersediaan dan kecukupan pangan
masyarakat.
Selain hal-hal di atas, kondisi georafis pun
mempengaruhi distribusi pangan.
5. Sosiologi
dan antroplogi
Pola konsumsi pangan masyarakat dipengaruhi oleh
kebiasaan dan budaya setempat.
6. Politik
Kondisi politik mempengaruhi kestabilan suatu
negara dan wilayah. Demikian pula dengan aktifitas masyarakat termasuk
aktifitas produksi dan distribusi pangan.
Dinding Sel Bakteri
PENDAHULUAN

PEMBAHASAN



Berikut
tabel perbedaan kandungan
antara dinding sel bakteri gram positif dengan dinding sel bakteri gram
negatif:

 1. Bakteri tanpa dinding sel
1. Bakteri tanpa dinding sel

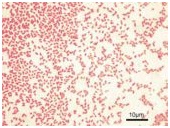

Bakteri merupakan mikroorganisme bersel satu yang
memiliki carakteristik yang bervariasi yang memungkinkan klasifikasinya. Satu
skema klasifikasi utama adalah berdasarkan pewarnaan Gram. Pada prosedur ini
bakteri dibunuh dengan pemanasan kemudian diwarnai dengan purple dye crystal
violet dan iodium. Kombinasi ini membentuk kompleks celupan pada dinding sel
bakteri. Perlakuan pewarnaan bakteri dengan sebuah dekolorisasi seperti etanol
akan mencuci kompleks dari beberapa bakteri yang membedakan dari yang lain.
Bakteri yang menahan kompleks kristal violet iodium terlihat ungu dan disebut
“Gram positif”. Bakteri yang yang kehilangan kompleks celupan dapat diwarnai
dengan celupan merah saffranin, terlihat merah dan disebut “Gram negatif”. Dasar
dari reaksi Gram terjadi karena struktur diding sel yang akan dijelaskan dalam
paper berikut.
Dalam paper ini juga akan dibahas tentang Bakteri
penyebab TBC yang merupakan bakteri tahan asam yang memliki dinding sel yang
unik. Bakteri ini pun dapat memiliki resistensi terhadap beberapa obat anti
TBC.

PEMBAHASAN
Dinding Sel Bakteri
Berat
dinding sel mencapai 40% berat kering sel bakteri. Dinding sel pada bakteri
tersusun atas peptidoglikan berbeda dengan dinding sel
tumbuhan (selulosa) atau dinding sel jamur (kitin).


{kind=link}
- N-asetil glukosamin
- Asam N-asetil muramat
- Peptida yang disusun empat atau lima asam amino, yaitu L-alanin, D-alanin, asam D-glutamat dan lisin atau asam diaminopimelat
- Komponen kimia lain seperti asam teikoat, protein, polisakarida, lipoprotein dan lipoposakarida yang terikat kuat pada peptidoglikan
Dinding sel
pada bakteri berupa struktur kaku yang terletak di sebelah luar membran sel.
Dinding sel berfungsi untuk :
- memberi bentuk pada sel
- memberi perlindungan
- berperan dalam reproduksi sel
- mengatur pertukaran zat dari dalam dan keluar sel. Dalam fungsinya membantu pertukaran zat; air, ion-ion dan molekul kecil dapat melintas dengan bebas melalui pori-pori kecil dalam dinding sel. Molekul besar seperti protein dan asam nukleat tidak dapat melalui pori-pori dengan bebas.
Kelompok
bakteri ada yang memiliki dinding sel
dan ada yang tidak. Bakteri yang berdinding sel sel dapat dibedakan berdasarkan
hasil pengecatan Gram
Perbedaan
utma dari dinding sel bakteri gram positif dengan dinding bakteri gram negatif
adalah: pada bakteri gram positif tersusun atas lapisan peptidoglikan relatif
tebal, dikelilingi lapisan teichoic acid dan pada beberapa species mempunyai
lapisan polisakarida, sedangkan dinding sel bakteri gram negatif mempunyai
lapisan peptidoglikan relatip tipis, dikelilingi lapisan lipoprotein, lipopolisakarida, fosfolipid
dan beberapa protein.
Peptidoglikan
(murein) yaitu susunan yang terdiri dari polimer besar dan terbuat dari
N–asetil glukosamin dan asam N–asetil muramat yang saling berikatan silang
(cross linking) dengan ikatan kovalen.

Beberapa
kelompok prokariota, secara umum disebut archaebacteria, tidak memiliki lapisan
peptidoglikan. Beberapa memiliki polimer
yang sama mengandung gula N-acetyl. Pada archaebacteria lain juga terdapat
lapisan protein.
Sel selama
mensintesis peptidoglikan memerlukan ensim hidrolase dan sintetase. Untuk
menjaga sintesis supaya normal, kegiatan kedua ensim ini harus seimbang satu
sama lain. Bio-sintesis peptidoglikan berlangsung dalam beberapa stadium dan
antibiotik pengganggu sintesis peptidoglikan aktif pada stadium yang berlainan.
Sikloserin terutama menghambat ensim racemase dan sintetase yang berperan dalam
pembentukan dipeptida. Vankomisin bekerja pada stadium kedua diikuti oleh
basitrasin, ristosetin dan diakhiri oleh penisilin dan sefalosporin yaitu menghambat
transpeptidase.
Diaminopimelic
acid merupakan elemen unik pada dinding sel prokariotik. Bakteri mutan yang
dihambat di tempat sebelum biosintesis zat ini, dapat tumbuh normal apabila
dalam medianya tersedia. Jika hanya L-lisin,
mereka mengalami lisis, karena pertumbuhan berjalan terus namun tidak mampu
membuat dinding sel peptidoglikan baru.
Banyak
spesies bakteri gram-negatif yang bersifat patogen, yang berarti mereka
berbahaya bagi organisme inang. Sifat patogen ini umumnya berkaitan dengan
komponen tertentu pada dinding sel gram-negatif, terutama lapisan lipopolisakarida (dikenal
juga dengan LPS atau endotoksin).
Antibiotik yang menyebabkan gangguan sintesis
lapisan ini aktivitasnya akan lebih nyata pada bakteri gram positif. Aktivitas
penghambatan atau membinasakan hanya dilakukan selama pertumbuhan sel dan
aktivitasnya dapat ditiadakan dengan menaikkan tekanan osmotik media untuk
mencegah pecahnya sel. Bakteri tertentu seperti mikobakteriadan halobakteria
mempunyai peptidoglikan relatip sedikit, sehingga kurang terpengaruh oleh
antibiotik
1. Bakteri tanpa dinding sel- sering dianggap virus (ukuran terlalu kecil dan tidak adanya dinding sel). Tetapi memiliki struktur seperti prokariot berupa ribosom, DNA dan RNA
- dapat hidup tanpa dinding sel karena hidup pada habitat dengan tekanan osmotik yang terlindung seperti pada sel tubuh hewan
- Contoh : Mycoplasma pneumoniae (bakteri patogen pada hewan dan manusia)
- Kebanyakan Mycoplasma memiliki sterol pada membran yang kemungkinan berfungsi untuk menambah kekuatan dan kekakuan membran seperti pada membran sel eukariot
{kind=link}
- Memiliki lapisan peptidoglikan tebal berupa asam teikoat
- Dinding sel yang tebal tersebut menyerap kristal violet saat pewarnaan Gram sehingga berwarna ungu/biru
- Mengandung lebih sedikit asam amino
- Contoh : Bacillus thuringiensis
3. Gram Negatif
- Memiliki lapisan peptidoglikan yang lebih tipis
- Memiliki kandungan lipid lebih tinggi
- Memiliki membran luar yang melindungi dari lingkungan yang tidak menguntungkan
- Memiliki lipoposakarida (LPS) sebagai materi endotoksin yang banyak dimiliki bakteri patogen
- Terdapat ruang periplasma yang berisi air, nutrien, hasil sekresi (enzim pencerna dan protein transport)
- Contoh : Pseudomonas

MYCOBACTERIUM
TUBERCULOSIS

Genus Mycobacterium
merupakan kelompok bakteri gram positip, berbentuk batang, berukuran lebih kecil
dibandingkan bakteri lainnya. Genus ini mempunyai karakteristik unik karena
dinding selnya kaya akan lipid dan lapisan tebal peptidoglikan yang mengandung
arabinogalaktan, lipoarabinomanan dan asam mikolat. Asam mikolat tidak biasa
dijumpai pada bakteri dan hanya dijumpai pada dinding sel Mycobacterium dan
Corynebacterium. Mycobacterium tuberculosis dibedakan dari sebagian besar
bakteri dan mikobakteri lainnya karena bersifat patogen dan dapat berkembang
biak dalam sel fagosit hewan dan manusia. Pertumbuhan Mycobacterium
tuberculosis relatif lambat dibandingkan mycobakteri lainnya.
Mycobacterium
tuberculosis tidak menghasilkan endo-toksin maupun eksotoksin. Bagian selubung Mycobacterium
tuberculosis mempunyai sifat pertahanan khusus terhadap proses
miko-bakterisidal sel hospes. Dinding sel yang kaya lipid akan melindungi
mikobakteri dari proses fagolisosom, hal ini dapat menerangkan mengapa
mikobakteri dapat hidup pada makrofag normal yang tidak teraktivasi.
Selain
bersifat patogen Mycobacterium tuberculosis dapat berfungsi sebagai ajuvan
yaitu komponen bakteri yang dapat
meningkat-kan respon imun sel T dan sel B apabila dicampur dengan antigen
terlarut. Ajuvan yang banyak digunakan dalam laboratorium adalah Freund's
ajuvan yang terdiri dari Mycobacterium tuberculosis yang telah dimatikan dan
disupensikan dalam minyak kemudian diemulsikan dengan antigen terlarut.
Mekanisme kerja obat antituberkulosis


OAT yang akan
dibahas dalam makalah ini adalah INH, RIF, PZA, EB dan SM. INH diduga bekerja
dengan menghambat sintesis asam mikolat, komponen utama dinding sel
Mycobacterium. RIF merupakan
bakterisid yang bekerja menghambat sintesis asam nukleat yaitu sintesis RNA transkripsi
dengan mengi-kat subunit β RNA polimerase. PZA merupakan analog struktur
nikotinamid yang membunuh basil tuberkel semidorman dalam keadaan asam. Dalam
keadaan asam, basil tuberkel menghasilkan pirazinamidase, suatu enzim yang
merubah PZA menjadi asam pirazinoat, yang berfungsi sebagai antibakteri. EB
mengganggu metabolisme karbohidrat, sedangkan SM membunuh bakteri dengan
mengganggu sintesis prote-in, translasi, dengan berikatan pada 16s rRNA.
Mekanisme tingkat molekul resistensi terhadap obat
antituberculosis isoniazid (INH)
INH
merupakan antituberkulosis yang paling efektif baik untuk pengobatan maupun
untuk pencegahan penyakit TB karena M. tuberculosis sangat sensitif terhadap
INH. Galur resisten INH seringkali muncul dengan frekuensi kurang lebih 90%.
Resistensi terha-dap INH disebabkan oleh mutasi pada salah satu dari gen katG,
inhA atau ahpC.
Setelah
masuk ke dalam sel, INH diubah menjadi bentuk aktifnya oleh enzim
katalase-peroksidase (KatG) yang dikode oleh gen katG. Diduga, INH yang aktif
berupa bentuk teroksidasinya. Enzim katG
meru-pakan satu-satunya enzim yang mengaktivasi INH, oleh karena itu hilangnya
aktivitas KatG akibat mutasi pada gen katG mengakibatkan M. tuberculosis
resisten terhadap INH. Resistensi terhadap INH yang paling banyak terjadi
adalah akibat mutasi pada gen katG. Yang banyak dilaporkan adalah mutasi pada
kodon 315 (Ser → Thr) dan kodon 463 (Arg → Leu) Selain itu, juga ditemukan
mutasi pada kodon 304 (Ile → Val) dan delesi parsial, di samping pada kodon 128
(Arg → Gln) dan kodon 291 (Ala → Pro), delesi sempurna pada gen katG dan
perubahan pada urutan peregulasi inhA yaitu substitusi C209T.
INH dalam
bentuk aktif menghambat enzim enoil-ACP reduktase, InhA (dikode gen inhA),
suatu enzim yang mengkatalisis tahap awal sintesis asam mikolat. Reaksi yang
dikatalisis oleh KatG terhadap INH diduga meng-hasilkan spesi elektrofil yang
dapat bereaksi dengan molekul sasaran dalam sel
Mycobacterium seperti InhA. Penelitian dengan pendekatan struktur menun-jukkan
bahwa INH-aktif bereaksi dengan NAD(H), suatu kofaktor yang terikat pada InhA
kemudian membentuk ikatan kovalen INH-NAD. Kepekaan terhadap INH disebabkan
karena penggabungan INH-NAD sehingga menghambat aktivitas enzimatik InhA.
Mutasi pada inhA juga menyebabkan Mycobacterium menjadi resisten terhadap
antituberkulosis lain yang struktur kimianya mirip INH yaitu etionamid. Mutasi
pada gen inhA belum banyak dilaporkan, namun telah ditemukan mutasi pada kodon
94 (Ser → Ala) dan daerah regulasi, yaitu posisi 209 C→T.
Sekitar 16%
isolat klinik yang resisten terhadap INH menunjukkan mutasi pada gen ahpC yang
mengkode AhpC, suatu enzim alkil hidroperok-sidase yang berfungsi sebagai
komponen reduktase antioksidan. Jika gen katG termutasi maka ekspresi ahpC
mening-kat untuk mengatasi hilangnya fungsi KatG. Mutasi yang bertanggungjawab
terhadap peningkatan ekspresi gen ahpC
adalah pada daerah berukuran 105 pasangan basa yang berlokasi di antara
oxyR-ahpC, biasanya terjadi pada frekuensi rendah berupa transisi G-C men-jadi
A-T.
Rifampin (RIF)
Terjadinya
resistensi terhadap RIF mencapai 95, umumnya terjadi akibat mutasi pada gen
rpoB yang mengkode subunit β RNA polimerase, komponen penting dalam proses
transkripsi. RIF terikat secara
spesifik pada subunit β RNA polimerase sehingga transkripsi terhambat. Berbagai
mutasi pada gen rpoB telah diketahui bertanggungjawab terhadap resistensi RIF,
terbanyak terjadi pada kodon 526 (His → Asp) dan kodon 531 (Ser → Leu). Mutasi
lain yang menyebabkan perubahan asam amino terjadi pada kodon 518 (Asn → Ser),
kodon 513 (Gln → Leu) dan delesi sembilan nukleotida. Di Afrika Selatan
ditemukan gen rpoB M. tuberculosis dengan kodon 531 termutasi (Ser →Trp). Di
Vancouver, Mexico City dan New Delhi, isolat M. tuberculosis resisten RIF
banyak yang mengalami mutasi pada kodon 526 (His → Arg), kodon 531 (Ser → Leu),
dan kodon 516 (Asp → Val) (Sharma, M, 2000), mutasi yang berupa misens atau
mutasi ganda. Ditemukan adanya mutasi gen katG pada kodon 505 (Phe → Leu), 511
(Leu → Pro), 516 (Asp → Ala), 525 (Thr → Asn), 526 (His → Tyr dan His → Leu),
531 (Ser → Trp) dan 533 (Leu → Pro). Mutasi delesi juga dilaporkan terjadi pada
posisi 199 – 207, yaitu hilangnya urutan ATGGACCAG, yang menyebabkan tiga asam amino
hilang, yaitu Met, Asp dan Gln dan pada kodon 354 terjadi delesi GGG, yang
menyebabkan kehilangan asam amino Gly. Selain mutasi tersebut, ditemukan juga
mutasi pada gen rpoB yang terletak di luar posisi yang biasanya terjadi yaitu
pada kodon 395 (Arg → Gln), 232 (His → Tyr), 221 (Ser → Leu), 202 (Asp → Tyr),
202-203 (Asp → Phe), 91 (Met → Leu), 227-228 (Leu → Ser), dan 349-351 (Gln →
Ser) (Schilke, K., 1999).
Pirazinamid (PZA)
PZA yang
masuk ke dalam M. tuberculosis akan diubah menjadi bentuk aktif oleh enzim
pirazinamidase (PZAase). Enzim PZAase dikode oleh gen pncA dan mutasi pada gen
pncA menyebabkan aktivitas PZAase hilang sehingga M. tuberculosis resistensi
terhadap PZA. Pada sejumlah isolat resisten PZA ditemukan mutasi berupa
substitusi nukleotida, insersi, delesi, substitusi asam amino atau pergeseran
kerangka baca, di antaranya pada kodon 118 Asn → Thr, insersi CG pada posisi 501,
insersi CC pada posisi 403, delesi 8 pb pada kodon start, kodon 54 (Pro → Thr),
insersi AG pada posisi 368, kodon 41 (Tyr → His), kodon 88 (Ser → stop kodon)
dan insersi A pada posisi 301. Pada 70 isolat resisten PZA yang dikarakterisasi
dalam suatu penelitian ditemukan 68 galur resisten tidak mengalami mutasi pada
gen pncA. Kenyataan ini membuka kesempatan untuk menemukan gen selain pncA yang
bertanggung jawab terhadap resistensi PZA. Dua galur lainnya tidak mempunyai
aktivitas PZAase yang didu-ga berhubungan dengan mutasi misens, salah satunya
mengalami mutasi gen pncA pada kodon 82 dan yang lain pada kodon 171 (Ala →
Val). Selain itu ditemukan juga dua mutasi lain yang mengakibatkan substitusi
fenilalanin oleh glisin pada posisi 80 (mutasi pada posisi 241, T → G), dan
substitusi alanin oleh prolin pada posisi 171 (mutasi pada posisi 511, G → C).
Etambutol (EB)
Resistensi
terhadap EB berkaitan dengan mutasi yang terjadi pada gen embB pengkode
arabinosiltransferase yang terlibat dalam
biosintesis arabinan, suatu kompo-nen arabinogalaktan pada dinding sel. Mutasi
pada gen embB dapat menghambat polimerisasi dinding sel arabinan dan
menyebabkan akumulasi karier lipid dekaprenol fosfoarabinosa. Diduga bahwa obat
meng-ganggu transfer arabinosa pada aseptor dinding sel.
Isolat M.
tuberculosis resisten paling umum mengala-mi mutasi pada gen embB pada kodon
306 atau 406 dimana terjadi substitusi asam amino. Pada kodon 306 dilaporkan
adanya substitusi of menjadi Dr, ug atau Rer. Selain itu dilaporkan juga adanya
mutasi pada kodon 285, 330 dan 630. Baik substitusi tunggal
misalnya pada kodon 313 (sis → tan) maupun substitusi ganda yaitu pada kodon
319 (Tyr → Cys) dan pada kodon 328 (Asp → Tyr). Mutasi lain menga-kibatkan
gen-gen pada operon iniBAC, operon yang terdiri atas gen iniA, iniB dan iniC,
diekspresi lebih tinggi bila M. tuberculosis dipaparkan terhadap etam-butol
secara in vitro.
Streptomisin (SM)
Mutasi yang
menyertai resistensi SM diketahui berhu-bungan dengan gen 16S rRNA (rrs) dan
gen protein ribosom S12 (rpsL), yang paling banyak mutasi pada gen rpsL dan
umumnya terjadi pada kodon 43. 54% isolat resisten SM mengalami mutasi misens
sehingga mengakibatkan substitusi asam amino pada kodon 43 (Lys → Arg). Mutasi
pada kodon 88 gen rpsL juga ditemukan. Mutasi pada gen rrs terjadi pada dua
daerah yaitu di sekitar nukleotida 530 dan 951 serta ditemukan trans-versi A →
T pada posisi 513. Ditemukan isolat resisten SM yang mengalami substitusi C865
→ G pada loop 912 gen rrs dan tidak ditemukan mutasi pada loop 530.
Konsekuensi mutasi terhadap aktivitas enzim
Mutasi dapat
disebabkan oleh perubahan nukleotida pada titik tertentu (mutasi titik),
hilangnya nukleotida baru pada satu titik
(delesi mikro) atau pada fragmen besar DNA (delesi makro). Selain dapat terjadi
pada daerah pengkode, mutasi dapat juga terjadi pada daerah yang bertanggung
jawab terhadap regulasi ekspresi OAT sasaran atau enzim pengaktivasi OAT,
misalnya pada promotor. Mutasi pada daerah promotor menga-kibatkan transkripsi tidak
terjadi atau turun sehingga enzim tidak dapat atau hanya sedikit disintesis.
Mutasi pada
daerah pengkode dapat menyebabkan substitusi asam amino pada protein dengan
ukuran normal atau menghasilkan protein yang lebih pendek. Substitusi asam
amino dengan sifat berbeda dapat menyebabkan protein kehilangan aktivitas
enzimatis atau kehilangan aktivitas pengikatan. Beberapa contoh substitusi asam
amino yang dapat menyebabkan kehila-ngan atau penurunan aktivitas enzimatis
terjadi pada enzim KatG dan PZAase. KatG mutan mengalami perubahan Arg463 →
Leu, dimana Arg yang bersifat basa, dan berukuran kecil menjadi Leu yang
bersifat netral, hidrofob dan berukuran besar. Jadi perubahan ukuran dan muatan
asam amino ini sangat bermakna sehingga KatG kehilangan aktivitas enzimatisnya.
Contoh lain adalah pada enzim ce ase mutan yang mengalami perubahan
Tyr41 → His, dimana kedua asam amino tersebut bersifat polar, namun telah
terjadi perubahan muatan (Tyr bersifat netral, sedangkan His bersifat basa)
Hilangnya aktivitas enzim juga dapat disebabkan oleh mutasi nonsens yang
menyebabkan terjadi kodon stop pada Ser88, sehingga enzim in ase yang
dihasilkan menjadi lebih pendek dan tidak mempunyai aktivitas.
Substitusi
asam amino juga dapat menghilangkan aktivitas pengikatan. Sebagai contoh
terjadi pada enzim InhA, dimana terjadi substitusi Ser94 → Ala. Walau-pun kedua
asam amino tersebut bersifat netral dan berukuran sangat kecil, tetapi terjadi
perubahan polari-tas karena serin bersifat polar (hidrofil), sedangkan alanin
bersifat tidak polar (hidrofob). Enzim InhA mutan tidak dapat diinhibisi oleh
INH-NAD. Kegaga-lan pengikatan juga ditunjukkan oleh dua mutasi pada gen rpoB
yang menghasilkan substitusi His → Asp pada posisi 526 dan Ser → Leu pada
posisi 531. Perubahan His menjadi Asp tidak merubah sifat kepolaran dan
hidrofilitasnya, melainkan merubah sifat keasamannya karena His bersifat basa,
sedangkan Asp bersifat asam. Hal ini berbeda pada mutasi yang menyebabkan
substitusi Ser531 → Leu karena kedua asam amino tersebut bersifat netral dan
hidrofob, namun kepolarannya berbeda (Ser bersifat polar, sedangkan Leu
bersifat tidak polar). Walaupun perubahan sifat asam amino pada kedua RpoB
mutan berbeda, tetapi fenotip yang tampak adalah sama, yaitu keduanya
menunjukkan aktivitas pengikatan yang menurun terhadap rifampisin. Substitusi
asam amino pada enzim EmbB juga menyebabkan afinitas pengika-tan terhadap
ethambutol juga hilang atau turun, yaitu perubahan Asp328 → Tyr, dimana
kepolaran kedua asam amino tersebut tidak berubah, namun muatannya berubah (Asp
bersifat asam, sedangkan Tyr bersifat netral).
Spain, T
Resistensi
terhadap banyak OAT pada M. tuberculosis terjadi akibat mutasi yang tidak
saling bergantung (independent mutation) pada lebih dari satu gen pengkode OAT
dan atau gen pengkode enzim pengaktivasi prazat OAT. Mutasi
yang terjadi dapat berupa substitusi asam amino dengan perubahan sifat atau
struktur asam amino, perubahan kodon menjadi kodon stop sehingga enzim yang
dihasilkan berukuran lebih pendek, perubahan kerangka baca akibat hilangnya
atau penambahan nukleotida tertentu. Mutasi ini mengakibatkan protein target
atau enzim pengaktivasi menjadi hilang aktivitas enzimatiknya atau aktivitas
pengikatannya.
Langganan:
Komentar (Atom)